introduction

私たちの体は、様々な「かたち」や「構造」を持った組織・器官で構成されています。さらに、そこには連動した固有の「機能」が備わっています。つまり、脳、心臓、血管、皮膚などの様々な機能を持つパーツをうまく組み合わせることで、からだをつくることが理論的には可能です。さらにスケールを変えて見てみると、組織・器官は機能的最小単位である“細胞”が集合することでできあがっています。組織によって参加している細胞の種類・割合は様々であり、細胞の組み合わせ方の違いだけでその多様性を生み出していることは非常に驚くべきことです。そこでは、集合した細胞同士が常にコミュニケーションをとることでお互いのふるまいを巧妙に制御し、それが繰り返されることで秩序ある集合体となっていく、私たちの想像を超えた世界が展開されています。
私たちは、なぜ細胞が集合すると自律的に「かたち」、「構造」そして「機能」が生じ得るのか、そのしくみに迫ることを大きな研究目標の一つとしています。脳や心臓といった最終形は違っても、細胞の集合体であること自体は同じですのでそれを生じさせる普遍的なしくみがそこには存在するはずです(図1)。一方、脳と心臓はかたちも機能も違うわけですから、それぞれに何か特別なしくみも存在するはずです。理解の対象が極めて複雑ですので、それぞれの側面をしっかりと切り分けて考えていくことが肝要であると思っています。組織や器官をつくり・維持するための根幹のしくみ(設計図)が分かれば、それらが破綻した状態である病気のしくみがよりよく理解できてきますし、それを基にして新しい治療法の開発にも繋がってきます。特に、からだができる基本設計図が分かってくれば、ある程度自在に体そのものやその一部をつくる次世代の再生医学も夢ではなくなると考えます。つまり、私たちの2つ目の大きな研究目標は、自身で得た科学を医学に役立てることです。
実際には、私たちは血管を対象とした研究を行っています。血管は、私たちの体のほぼどこにでも存在する主要な器官の一つです。最も主要な血管機能として、私たちの全身の組織に酸素や栄養を運搬する管としての役割があげられます。特記すべきことは、血液を効率良く組織に灌流するために、非常に合理的な血管配置と血管径が適宜設定されていることです。このような血管ネットワークとしての合理性は、血管がつくられたり壊されたりすること、そして血管径が大きくなったり小さくなったりすることで構造的かつ機能的に築き上げられています。血管の主な構成細胞は、血管内皮細胞と壁細胞/平滑筋細胞です(動脈では、組織の支持細胞として線維芽細胞も大きな構成細胞となってきます)。驚くことに、このたった2種類の細胞が集合することで、管腔や枝分かれネットワークなどの複雑な「かたち」や「構造」をつくり、さらにその形態自体が血管としての「機能」に連動しています。私たちは、細胞が集合するとなぜ自律的に「かたち」、「構造」そして「機能」が生じるのかという一般的な問いを理解するため、血管は非常に良いモデルになると考えています。

ではどのようなアプローチで研究をすれば、私たちの問いに答えることができるのでしょうか? もちろん私たち自身もその正解はわかりませんが、少なくとも問題設定に沿ってアプローチ法自体も常に改良・開発していく研究展開が必要ではないかと考えています。単なる多細胞体が血管のような複雑な機能性構造体になる過程では、細胞の分裂や変形、そして多細胞運動など、多くの細胞現象が起こっています。したがって、この過程を隈なく観るということが研究の大きな軸の一つになるはずです。つまり、イメージングです。次に、観た現象を何らかのかたち(指標)で定量化する必要があります。例えば、細胞の動きは速度という指標で定量的にその一部を現すことができます。現象を定量化できれば、分子動態や細胞動態、形態変化といった違う現象レベルを繋ぐ関係性を統計学的に推測することが可能となります。さらに、観ることができた複雑現象を動かしている、観ることができないしくみをも理解する必要があります。ここでは数理生物学の力を借りてそれを予測します。予測できたしくみは、非常に単純な実験系を用いることで再現し、それにより予測したしくみの正当性を検証します。私たちは、現象をできるだけ単純化したかたちでスタートし徐々に複雑化していくことで理解する、構成的実験アプローチを2つ目の実験軸として研究を行っています(図2)。
ここまで、私たちはどのようなスタンスで研究を行っているかを長々と記載してきました。私たちの研究に少しでも興味が湧いた方は、以下の研究概要へと読み進みください。以降は、より専門的な内容になってきます。
Research outline
研究概要
血管は、私たちの体ほぼ全てに存在し、組織に酸素や栄養を供給するために必須な管状の器官です。組織に効率良く血液を運ぶために、血管は極めて合理的なネットワーク構造を呈しています。その全体構造は、組織の状況(個体発生、虚血組織、がん等)に応じて大きく変化し(動的)、また、組織の違い(心臓、網膜、脳等)によって大きく異なってきます(多様性)。私たちの研究の一つは、血管ネットワークを動的に構築するための主要な要素である出芽的血管新生とリモデリングのしくみを包括的に明らかにすることです。そのために、出芽的血管新生において、血管内皮細胞と血管壁細胞が協調し自己組織化的に樹状構造をつくるしくみを解きます(研究1)。さらにその理解に基づき、血管内腔からかかるずり応力などの物理的刺激が、出芽的血管新生動態やリモデリングによる血管成熟にどのように影響を与えているのか検討します(研究2)。また、組織依存的な血管ネットワーク構造の多様性にも焦点を当てた研究を行います(研究3)。ここでは、上で述べた血管内皮細胞と壁細胞による自発的な血管新生が、血管外の微小環境からどのような影響を受けているのか、そして、いかにその影響が構造の多様性へと帰結しているのかを問います。最終的には、血管形成や機能の恒常性破綻に起因する病態の理解や新規治療の開発に繋げることを目指しています。
(私たちの研究の基本理念は、‘はじめに’に記載してあります)
Research 1
研究1:血管新生動態を制御するしくみの解析
(1)血管新生動態
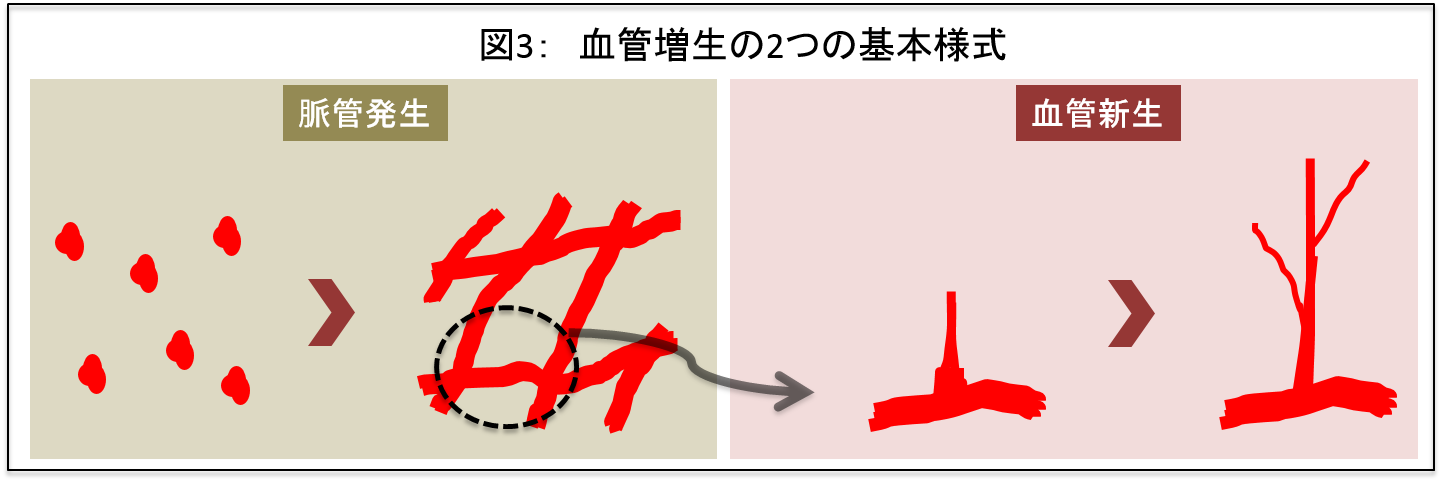
血管は、脈管発生と血管新生の大きな2つの過程により増生されます。脈管発生が血管の全くないところから初期血管網を形成する過程であるのに対して、血管新生は一度できた血管網から出芽的に血管が増生される現象を指します(図3)。私たちは、主に後者の血管新生に焦点を当てた研究を行っています。
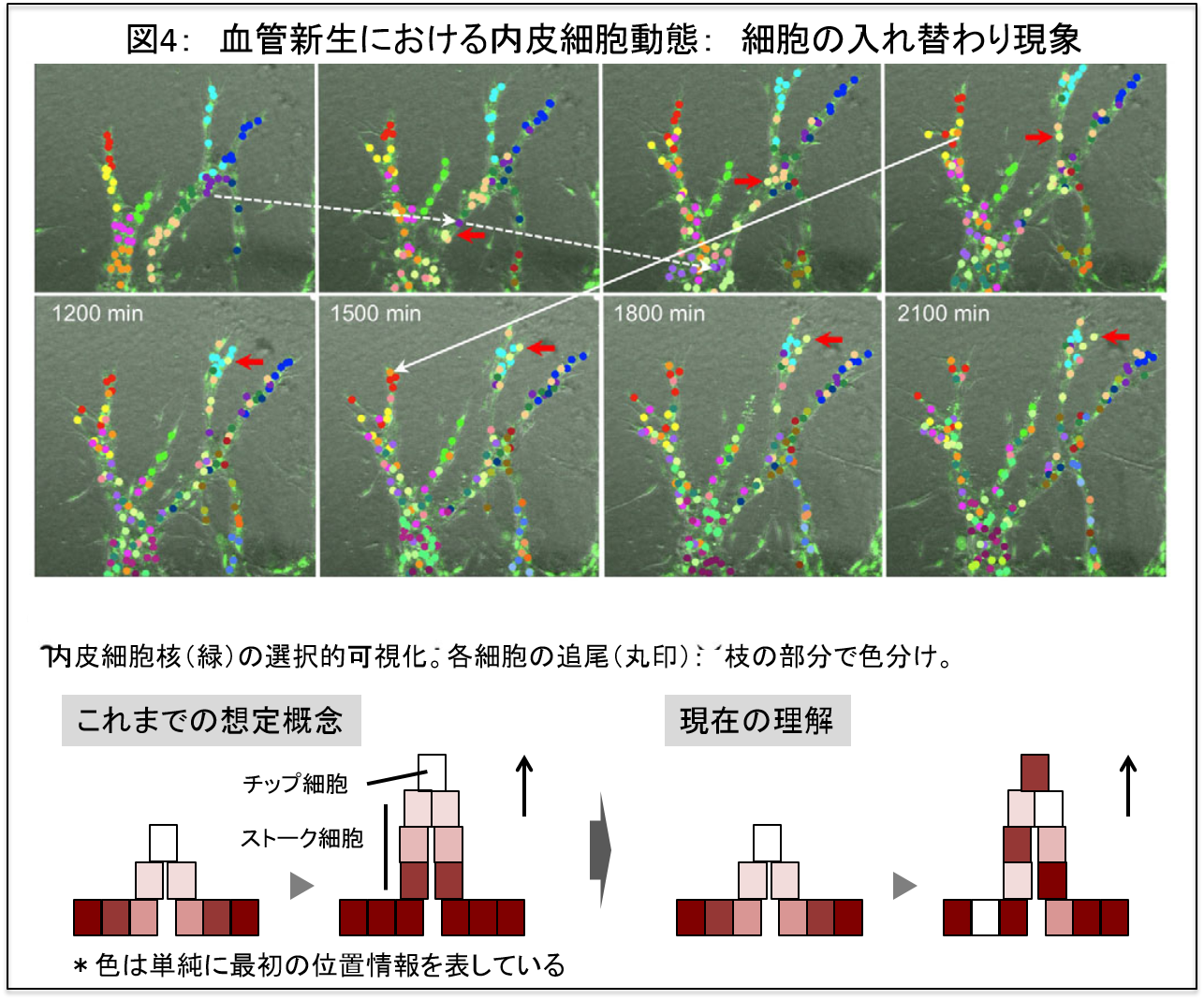
これまでに、血管新生をよく模すex vivo血管新生モデル(大動脈リングアッセイ)を用いて、血管新生の4次元動態をタイムラプス観察した結果、非常に興味深いことが分かってきました。血管の枝先端部分の成長には、内皮細胞の移動(集団的細胞運動)が重要であり、その移動はお互いに位置関係を常に入れ替える極めて動的で複雑な現象として捉えられてきました(図4)。特に、枝先端は先端細胞という特別な細胞が先導し茎細胞群がそれに後続する比較的静的な現象であると想定されていましたが、私達 (Development 2011) や他のグループ (Jakobsson et al. Nat Cell Biol 2010)の発見にて再考を迫られることになりました(図4)(これらのex vivoにおける細胞動態の知見は、マウス網膜血管新生においてin vivoでも確認)。現在では、細胞の入れ替わり現象は血管新生研究者には広く受け入れられています。
*他の参考文献:
・Development 138:4569-4583, 2011
・Curr Opin Cell Biol 24:188-193, 2012
(2)4次元イメージングと動態解析
これまで血管新生の際の細胞動態に関しては、ほぼ不明の状態でした。私たちは、上記知見の取得に先立って血管新生における多細胞動態を解析できる実験系を構築しました(図4、Development 2011)。血管新生においては、内皮細胞とそれを取り囲む壁細胞が集合体として血管構造をつくっていきますが、SYTO Green (Molecular probe)で核染色し顕微鏡の至適条件にて撮像することにより、内皮細胞核のみを生細胞で選択的に可視化することに成功しました。この技術の進歩により、同研究は飛躍的に進展しています。さらに、各細胞をImage JのプラグインソフトウエアであるMTrackJを用いて追尾し、時間軸で繋がった座標データを得ます。そのデータを基に、MATLABなどの数値解析ソフトを用いて、形態変化や細胞動態を模式化することや定量的に評価することを行ってきました。図5は、枝の伸長時の多細胞動態を2次元的に模式化したものです。このグラフを見ると、枝の伸長時の細胞動態変化を直感的に捉えることが可能となります。また、細胞動態を様々な指標で定量化することで、遺伝的介入を行った場合の細胞動態変化を評価でき、細胞現象と支配遺伝子といった異なるレベルでの事象を関連づけることが可能となりました。
(3)血管内皮細胞動態を制御するしくみの検討
ex vivo血管新生モデルのイメージングにより、血管新生的に血管形態をつくる時の多細胞動態が現象論的に明らかになってきました。私たちは、生体内の血管新生動態を総括的に理解するためには、先ず、ex vivo血管新生モデルで見られる血管内皮細胞と壁細胞が自発的に血管形態をつくる現象(自己組織化的)の背後に存在するしくみを十分理解する必要があると考えています。このしくみは、現象論的に十分には観察できませんので、数理生物学やコンピューターシュミレーションの力を借りることで統合的に解析しています。
これまでに、最も単純な血管新生要素である枝の伸長現象に関して検討を行った結果、内皮細胞動態制御機構として、細胞自律的および非自律的なしくみが少しずつ見えてきています。血管の伸長とその際の多細胞動態の大きな特徴は、個々の内皮細胞がそれぞれ確率的に運動するという単純なしくみだけでも十分説明できうることがわかってきました。さらに、数理モデルとゼブラフィッシュ胚を用いた実験にて先端細胞動態をより詳細に検討した結果、先端細胞の枝伸長方向への方向運動制御には直下の後続細胞動態が関与していることがわりました(Cell Reports, in press)。この論文では、数理生物学的解析において三浦岳博士(九州大学)と小林亮博士(広島大学)、マウス網膜の実験において植村明嘉博士(名古屋市立大学)、ゼブラフィッシュ胚を用いた解析において福原茂朋博士(国循)等、多数の共同研究者に支えられています。
現在、以下の問いに挑んでいます。
1) 内皮細胞運動を制御する内皮細胞間作用と分子実体
2) 内皮細胞運動を制御する壁—内皮細胞間作用と分子実体
3) 分岐形成メカニズム
4) マウス組織からの血管内皮・壁細胞の単離技術の確立
特に、1細胞におけるランダムに近い運動が、方向性運動、秩序ある多細胞運動、そして形態へと様々な細胞間作用を介してどのように制御されていくのか、構成的実験系を構築するところから研究をスタートさせています。
Research 2
研究2:血管新生制御における物理的因子の検討
研究1では、血管新生を内皮細胞と壁細胞が集合し枝状の構造をつくる現象として単純化して考えています。実際の血管新生では内部の管腔構造に血液が流れ、そこには血管壁に対して内腔からの垂直抗力(内腔圧)やせん断応力(ずり応力)といった物理的な力が加わっています。私たちは、血管新生による初期の血管網形成やリモデリングによる後期の血管成熟過程において、どのように物理的刺激が関わっているのか、もしくはいなかという問いに、細胞動態の観点から取り組んでいます。これまでに、ずり応力が血管の機能的拡張、退縮抑制などの恒常性維持や血管の成熟・分化に関わっていることがわかっていますが、その過程における細胞レベルでの事象はまだよく理解されていません。また、血管新生早期におけるずり応力の重要性は不明であり、さらに、内腔圧の関与に関してはほとんど検討されていないのが現状です。私たちは、物理的刺激を感知した内皮細胞や壁細胞が同種・異種細胞間で情報を伝達し、それが多細胞動態に反映されることで形態・機能形成に帰結していくしくみを知りたいと考えています。
そのためには生体内で起こる現象をそのまま観察することも必要ですが、それだけでは‘なぜ’というところには十分に切り込めませんので、生体内で複雑に起こっている事象をできるだけ単純化して解析できる構成的実験系を構築することが必須であると考えています。その大きな軸となるのが、微小流体デバイスを用いた実験系です。これを用いることで、流れや圧力といった物理量を実測もしくは推測しながら細胞動態を観察し、また、その操作による動態変化を解析することが可能となります。また、力を受ける側の細胞にも遺伝子操作などで介入することで、分子メカニズムにも迫ることができます。これまでに、微小流体デバイス内に内皮細胞と壁細胞による血管新生現象を再現し、その内腔に培養液を灌流することを可能としてきました。これらの多くの研究は、三浦岳博士(九州大学)と横川隆二博士(京都大学)との共同研究で行っており、CREST生命動態からの多くの補助により支えられています。
http://www.jst.go.jp/kisoken/crest/project/35/14532039.html
具体的には以下の研究を行っています。
1) 血管新生におけるずり応力と内腔圧の関与メカニズム− 壁細胞の役割
2) 流れによる動脈化メカニズムと誘導
Research 3
研究3:組織依存的な血管網構築メカニズム
研究1、2では、主に血管内皮細胞と壁細胞の集合体が持つ固有の形態形成能に着目した研究を行っています。さらに研究2では、流れを加えることにより生体内に一歩近づけたかたちの研究を行っていますが、各組織共通の血管新生のしくみにアプローチすることを主眼に置いています。ここでは、組織による血管網パターンの違いや構造の多様性が生まれるしくみに関して、特に血管新生とリモデリングに焦点をあてた研究を行います。私たちは、血管形成の共通のしくみが組織環境の違いにより修飾され組織特異性が生じるという基本概念にたち研究を進めています。
現在は、心筋内の血管新生に関する研究を行っています。虚血などの病的状態において心筋内血管新生は本当に起こっているのか、その根本的な問題は未だ未解決のままです。しかし、少なくとも発生期の心筋内では血管新生により血管が増生されることが示されています。心筋内微小環境は心筋細胞の収縮により常に拍動しています。つまり、血管が新生する場は常に空間的に大きく変化しており、そこには大きな物理的な力が働いています。私たちは、血管新生が起きるには一見過酷に思える心筋内微小組織において、環境から受ける物理的刺激にどう応答し、またはそれを回避しながら血管新生を行っているのかに非常に興味を持っています。ここでも、生体内への古典的解析アプローチに加えて、心筋組織培養による観察と心筋微小組織の再構築系も駆使して問いに向かうべく、その準備を進めています。
