(1)血管新生動態
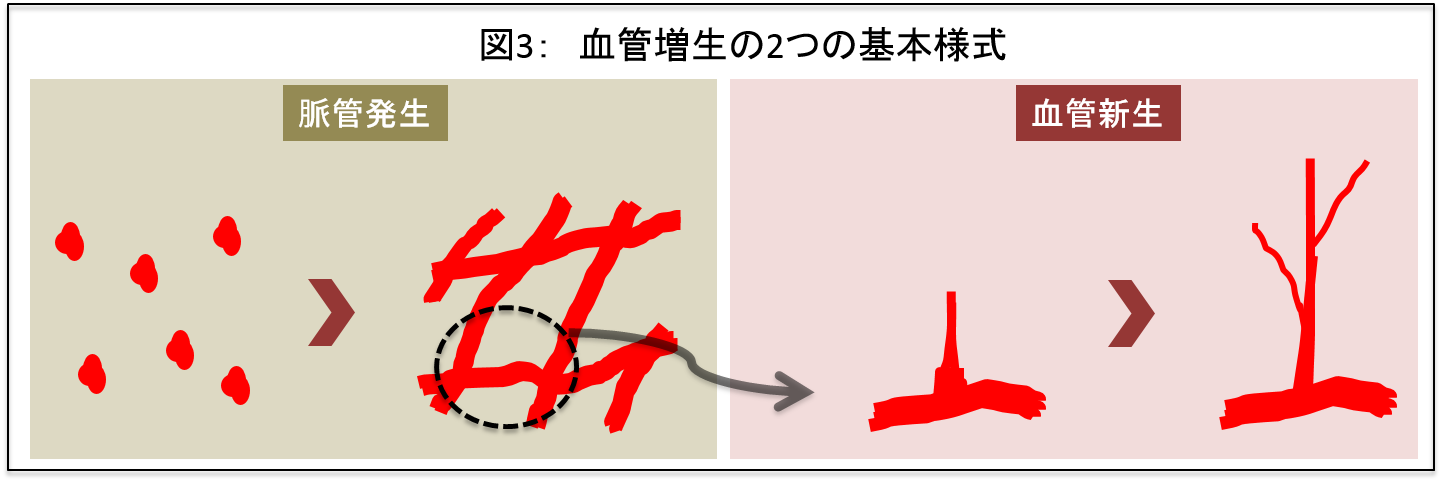
血管は、脈管発生と血管新生の大きな2つの過程により増生されます。脈管発生が血管の全くないところから初期血管網を形成する過程であるのに対して、血管新生は一度できた血管網から出芽的に血管が増生される現象を指します(図3)。私たちは、主に後者の血管新生に焦点を当てた研究を行っています。
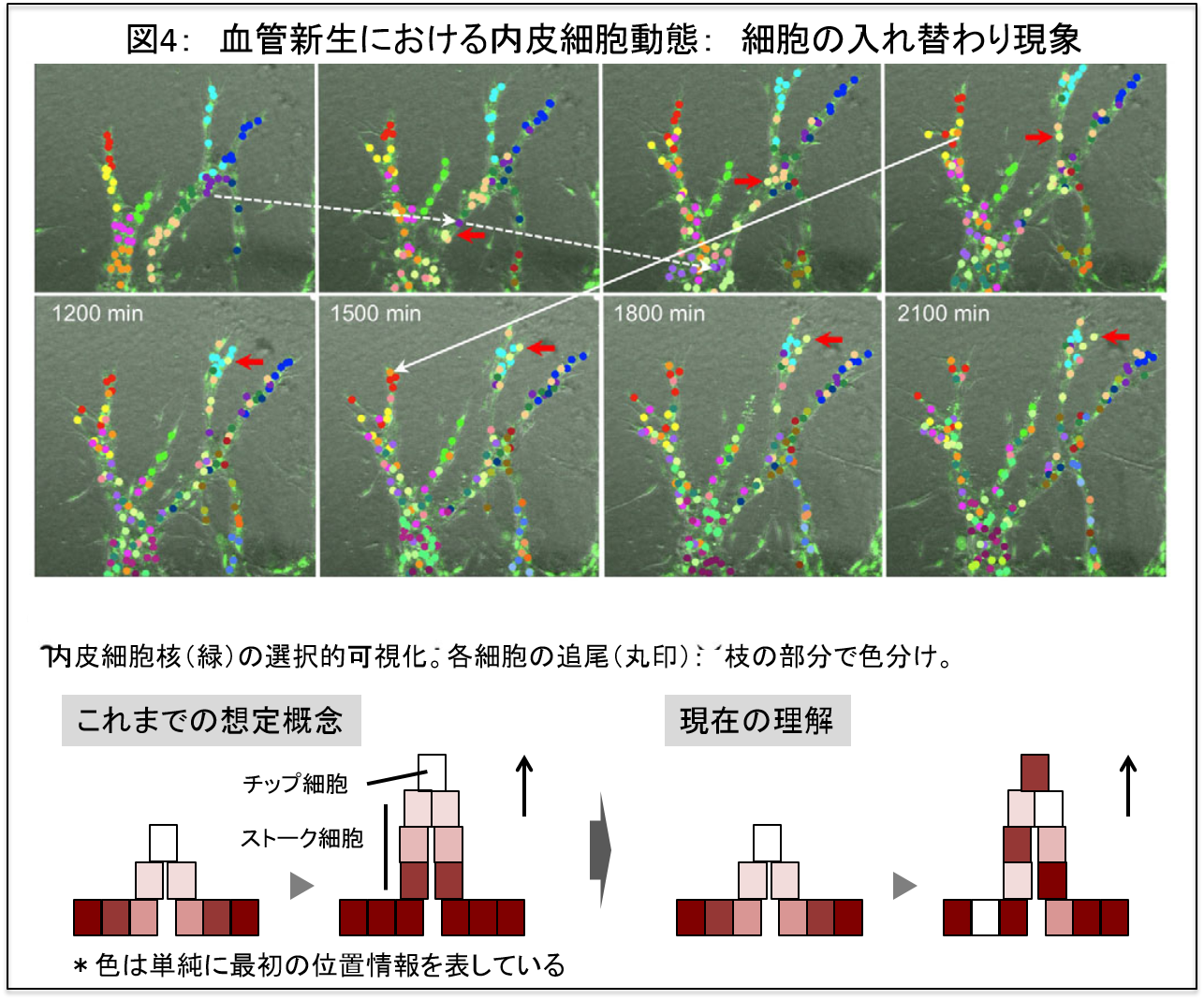
これまでに、血管新生をよく模すex vivo血管新生モデル(大動脈リングアッセイ)を用いて、血管新生の4次元動態をタイムラプス観察した結果、非常に興味深いことが分かってきました。血管の枝先端部分の成長には、内皮細胞の移動(集団的細胞運動)が重要であり、その移動はお互いに位置関係を常に入れ替える極めて動的で複雑な現象として捉えられてきました(図4)。特に、枝先端は先端細胞という特別な細胞が先導し茎細胞群がそれに後続する比較的静的な現象であると想定されていましたが、私達 (Development 2011) や他のグループ (Jakobsson et al. Nat Cell Biol 2010)の発見にて再考を迫られることになりました(図4)(これらのex vivoにおける細胞動態の知見は、マウス網膜血管新生においてin vivoでも確認)。現在では、細胞の入れ替わり現象は血管新生研究者には広く受け入れられています。
*他の参考文献:
・Development 138:4569-4583, 2011
・Curr Opin Cell Biol 24:188-193, 2012
(2)4次元イメージングと動態解析
これまで血管新生の際の細胞動態に関しては、ほぼ不明の状態でした。私たちは、上記知見の取得に先立って血管新生における多細胞動態を解析できる実験系を構築しました(図4、Development 2011)。血管新生においては、内皮細胞とそれを取り囲む壁細胞が集合体として血管構造をつくっていきますが、SYTO Green (Molecular probe)で核染色し顕微鏡の至適条件にて撮像することにより、内皮細胞核のみを生細胞で選択的に可視化することに成功しました。この技術の進歩により、同研究は飛躍的に進展しています。さらに、各細胞をImage JのプラグインソフトウエアであるMTrackJを用いて追尾し、時間軸で繋がった座標データを得ます。そのデータを基に、MATLABなどの数値解析ソフトを用いて、形態変化や細胞動態を模式化することや定量的に評価することを行ってきました。図5は、枝の伸長時の多細胞動態を2次元的に模式化したものです。このグラフを見ると、枝の伸長時の細胞動態変化を直感的に捉えることが可能となります。また、細胞動態を様々な指標で定量化することで、遺伝的介入を行った場合の細胞動態変化を評価でき、細胞現象と支配遺伝子といった異なるレベルでの事象を関連づけることが可能となりました。
(3)血管内皮細胞動態を制御するしくみの検討
ex vivo血管新生モデルのイメージングにより、血管新生的に血管形態をつくる時の多細胞動態が現象論的に明らかになってきました。私たちは、生体内の血管新生動態を総括的に理解するためには、先ず、ex vivo血管新生モデルで見られる血管内皮細胞と壁細胞が自発的に血管形態をつくる現象(自己組織化的)の背後に存在するしくみを十分理解する必要があると考えています。このしくみは、現象論的に十分には観察できませんので、数理生物学やコンピューターシュミレーションの力を借りることで統合的に解析しています。
これまでに、最も単純な血管新生要素である枝の伸長現象に関して検討を行った結果、内皮細胞動態制御機構として、細胞自律的および非自律的なしくみが少しずつ見えてきています。血管の伸長とその際の多細胞動態の大きな特徴は、個々の内皮細胞がそれぞれ確率的に運動するという単純なしくみだけでも十分説明できうることがわかってきました。さらに、数理モデルとゼブラフィッシュ胚を用いた実験にて先端細胞動態をより詳細に検討した結果、先端細胞の枝伸長方向への方向運動制御には直下の後続細胞動態が関与していることがわりました(Cell Reports, in press)。この論文では、数理生物学的解析において三浦岳博士(九州大学)と小林亮博士(広島大学)、マウス網膜の実験において植村明嘉博士(名古屋市立大学)、ゼブラフィッシュ胚を用いた解析において福原茂朋博士(国循)等、多数の共同研究者に支えられています。
現在、以下の問いに挑んでいます。
1) 内皮細胞運動を制御する内皮細胞間作用と分子実体
2) 内皮細胞運動を制御する壁—内皮細胞間作用と分子実体
3) 分岐形成メカニズム
4) マウス組織からの血管内皮・壁細胞の単離技術の確立
特に、1細胞におけるランダムに近い運動が、方向性運動、秩序ある多細胞運動、そして形態へと様々な細胞間作用を介してどのように制御されていくのか、構成的実験系を構築するところから研究をスタートさせています。